Trois ans après son opération, Camille croyait le sujet classé. La vésicule retirée, la convalescence passée, le médecin rassurant : « tout est rentré dans l’ordre ». Et c’est vrai la plupart du temps. Sauf certains matins. Une crampe, une envie pressante, et cette scène qu’elle connaît par cœur : si elle file aux toilettes, c’est liquide ; si elle parvient à attendre quelques minutes, l’envie reflue et la selle suivante est parfaitement normale. Entre deux épisodes, des semaines tranquilles. Puis un dîner un peu riche, un café avalé à jeun, une période de stress, et tout recommence. Ce qui l’angoisse, au fond, ce n’est pas la diarrhée c’est de ne jamais savoir quand.
Cette histoire est d’une banalité frappante. Et elle cesse d’être mystérieuse dès qu’on comprend ce que faisait, exactement, l’organe qu’on a retiré.
La vésicule n’était pas un sac, mais un chef d’orchestre
Le foie fabrique la bile en continu, jour et nuit, un demi-litre environ par jour. Cette bile contient les acides biliaires, de véritables détergents qui émulsifient les graisses pour permettre leur digestion. On imagine souvent la vésicule comme un simple réservoir. C’est réducteur : c’était surtout un organe de mise en forme du flux. Entre les repas, elle stockait la bile et la concentrait. Au moment où le gras arrivait dans l’intestin, elle se contractait et délivrait, d’un coup, une bouffée de bile concentrée, synchronisée avec le besoin.
Retirez-la, et ce n’est pas la bile qui manque, le foie continue d’en produire autant. C’est la synchronisation qui disparaît. La bile s’écoule désormais en goutte-à-goutte permanent, désynchronisée des repas, y compris à jeun, quand il n’y a rien à digérer. Imaginez un robinet qu’on ouvrait pile au moment du repas, remplacé par un filet qui coule en continu. La moyenne sur la journée est à peu près la même. Mais le timing, lui, a changé.
Un corps qui n’est pas « cassé », mais à réserve réduite
Voilà pourquoi « tout est revenu à la normale » est une demi-vérité. En moyenne, oui. Le côlon réabsorbe l’eau, le transit se stabilise, et chez une part notable des personnes opérées, les troubles des premières semaines rentrent dans l’ordre en quelques mois. Une minorité garde une gêne durable, mais pour la plupart, l’équilibre revient.
Sauf que la marge de sécurité, elle, a diminué. La vésicule était un tampon qui absorbait les à-coups. Sans elle, le système fonctionne près de son seuil : tant qu’on reste en dessous, le côlon gère sans problème ; mais une perturbation transitoire comme un repas gras copieux, le café à jeun, l’alcool, une grosse charge d’aliments fermentescibles, un pic de stress suffit à le faire basculer au-dessus. D’où l’épisode. Puis le système revient à son équilibre. C’est exactement ce modèle qui explique ces crises isolées surgissant des années après l’opération, sur un fond par ailleurs normal.
Le stress, d’ailleurs, n’est pas ici une formule polie pour « c’est dans la tête ». Il accélère directement la motricité du côlon : moins de temps pour réabsorber l’eau, donc des selles plus liquides. C’est un maillon physiologique réel de la chaîne, pas une cause imaginaire.
Ce déplacement de regard compte plus qu’il n’en a l’air. Passer de « mon corps est cassé et imprévisible » à « système stable, à réserve réduite, qui réagit à des déclencheurs identifiables » change l’expérience. Et tenir un simple journal déclencheurs ↔ épisodes transforme l’« imprévisible » en « conditionnellement prévisible ». Or ce qui angoisse n’est pas tant la diarrhée que l’imprévisibilité : rendre les épisodes lisibles, c’est déjà désamorcer une bonne part de l’anxiété.
« Si je me retiens, ça passe » : une question de vitesse, pas de panne
Le phénomène que décrit Camille mérite qu’on s’y arrête, car beaucoup le vivent sans l’expliquer. L’envie pressante correspond à une vague de contraction qui pousse le contenu vers le rectum. Aller à la selle pendant cette vague, c’est évacuer un contenu transporté trop vite, que le côlon n’a pas eu le temps de déshydrater : résultat, c’est liquide. Se retenir laisse la vague passer ; le rectum s’apaise, le côlon poursuit son travail de réabsorption, et la selle suivante est formée.
Autrement dit : c’est un problème de vitesse et de timing, pas une défaillance de l’organe. La capacité d’absorption est intacte. Et l’envie est souvent différable, savoir attendre quelques minutes est même une technique reconnue en rééducation de l’urgence. Comprendre cela, c’est déjà reprendre une part de contrôle sur ce qui semblait subi.
Le piège contre-intuitif : vouloir « soutenir la bile »
C’est ici que l’intuition trahit. Le réflexe spontané, y compris chez des personnes averties, est de se dire : « je n’ai plus de vésicule, je dois donc soutenir ma bile. » Et de se tourner vers les plantes réputées stimuler le foie et la bile : artichaut, boldo, radis noir, et autres « draineurs hépatiques ».
Pour le profil le plus fréquent, c’est exactement le contre-sens à éviter. Quand le problème vient d’un excès d’acides biliaires qui atteignent le côlon et y déclenchent une sécrétion d’eau, stimuler encore la bile revient à ajouter de l’irritant là où il y en a déjà trop. On aggrave ce qu’on croyait soulager. Et les plantes vendues comme « régulatrices » ne sont pas davantage un raccourci : sur ce terrain, aucune ne dispense d’avoir d’abord identifié le mécanisme.
Tout dépend en réalité du profil : une digestion difficile des graisses, une douleur de type spasme, une pullulation bactérienne dans l’intestin grêle ne se gèrent pas de la même façon. C’est précisément pourquoi il faut profiler avant d’agir, un travail qui gagne à être mené avec un·e praticien·ne formé·e, plutôt qu’à coups de réflexes intuitifs.
Ce qui aide vraiment, sans recette miracle
Quelques principes, valables pour beaucoup, sans se substituer à un avis personnalisé :
Structurer les repas plutôt que de les subir. De petites portions régulières, avec un gras réparti plutôt qu’un gros repas gras ponctuel, sollicitent moins un afflux de bile qu’on ne peut plus produire à la demande. À l’inverse, le régime ultra-pauvre en graisses est un faux ami : il n’est pas justifié et finit par nuire à la nutrition.
Les fibres solubles, flocons d’avoine, psyllium, forment un gel qui raffermit les selles et piège une partie des acides biliaires. C’est l’un des leviers les plus simples et les mieux compris.
Identifier ses propres déclencheurs avec un journal, on l’a vu, vaut souvent mieux qu’une longue liste d’interdits. Et travailler l’anticipation et le stress, respiration lente, pratiques de régulation, agit sur une cause réelle des épisodes, pas seulement sur le moral.
Quand il faut consulter
Comprendre ne dispense pas de vigilance. Certains signes sortent du cadre d’un simple inconfort fonctionnel et imposent un avis médical : un amaigrissement, du sang dans les selles, une fièvre, une coloration jaune de la peau ou des yeux, des réveils nocturnes répétés par la diarrhée, ou tout changement net et durable de vos habitudes.
Un point mérite d’être martelé : la diarrhée liée aux acides biliaires se traite efficacement. Un médicament prescrit par le médecin (un séquestrant des acides biliaires) peut transformer le quotidien. Personne ne devrait la subir des années sans qu’un diagnostic soit posé.
Pour aller plus loin
Camille n’a pas changé de corps. Elle a changé de récit : d’un organe « cassé » et capricieux à un système cohérent, à réserve réduite, dont elle a appris à lire les seuils. C’est souvent là que commence le mieux-être, non dans une plante miracle, mais dans la compréhension.
Pour les praticien·ne·s, ou les lecteurs et lectrices qui veulent entrer dans le détail du raisonnement, les différents profils cliniques, la façon de les distinguer par le questionnement, les axes d’accompagnement et leurs limites, ainsi qu’une bibliographie scientifique de référence — la fiche praticien complète est disponible en téléchargement libre ci-dessous.
Il y a une contradiction que je porte au quotidien et que je n’ai jamais vraiment réussi à classer. Je suis végane, et je suis opposé à l’expérimentation animale. Pas par sensiblerie : par cohérence. Si je refuse d’instrumentaliser un animal pour mon assiette et dans ma vie quotidienne, je vois mal au nom de quoi je l’accepterais pour produire une donnée. Et pourtant, quand je prépare un cours ou que j’argumente une stratégie, je me retrouve régulièrement à m’appuyer sur des études faites sur des animaux. Ça me met mal à l’aise. La question qui m’a tenu éveillé un soir tient en une phrase : présenter ces études fait-il de moi un soutien à ce que je condamne ?
Je veux prendre cette gêne au sérieux, sans la dissoudre dans une justification confortable, mais sans en faire non plus une paralysie.
Deux questions qu’on a tort de confondre
En y regardant de près, il y a deux questions emboîtées, et c’est leur confusion qui crée le malaise.
La première est rétrospective : que faire d’un savoir déjà produit ? La seconde est prospective : que soutenir pour l’avenir ?
Citer une étude répond à la première. Mon véganisme, mon refus de la vivisection, relèvent de la seconde. Rien n’oblige à ce que la réponse à l’une contamine l’autre. Le faux dilemme, c’est de croire que je n’ai le choix qu’entre présenter l’étude (donc cautionner) ou l’ignorer (donc rester pur). Il existe une troisième voie, et c’est elle que je vais défendre.
Démonter le mot « complicité »
« Complicité » recouvre en réalité trois choses très différentes.
La complicité causale. Ma citation provoque-t-elle de nouvelles expériences ? Pour une étude déjà publiée, l’effet est quasi nul. Le tort est consommé, irréversiblement. L’animal ne sera ni épargné par mon silence, ni de nouveau sacrifié par ma citation. C’est radicalement différent d’acheter un produit testé sur animaux, où mon achat est un signal de demande bien réel. Ici, la boucle de rétroaction directe n’existe pas.
La complicité expressive. Citer, est-ce endosser la méthode ? Non. Sauf à confondre des actes de parole distincts. Dire qu’un résultat est vrai et pertinent, ce n’est pas dire que la manière de l’obtenir était légitime, ni recommander de la répéter. Le savoir a une histoire que je n’endosse pas en bloc ; personne ne le fait.
La complicité systémique. C’est la seule qui me résiste vraiment. Une discipline où les modèles animaux sont massivement cités entretient leur prestige, donc leur centralité, donc, diffusément, sur le long terme, leur reconduction. C’est réel, mais c’est une rétroaction faible et collective, pas une responsabilité individuelle traçable. Il faut la nommer honnêtement, sans en faire le procès d’une simple citation.
L’objection que je garde intacte
Je pourrais m’arrêter là, soulagé. Je m’y refuse, parce qu’il existe un contre-argument solide que je ne veux pas écraser.
L’éthique a une dimension qui ne se réduit pas aux conséquences. Une communauté qui refuse par principe d’utiliser des données arrachées à la souffrance entretient une norme, et les normes finissent par façonner ce qui sera jugé acceptable demain. À cela s’ajoute un argument de dignité : utiliser la donnée, c’est traiter l’animal comme pur moyen, rétrospectivement encore ; refuser, c’est une manière de l’honorer comme plus qu’un instrument.
Si l’on tient ces deux raisons pour plus lourdes que tout le reste, l’abstention devient cohérente. Je le reconnais. Simplement, elle a un prix et ce prix est élevé.
L’ignorance désarme celui qui veut abolir
Voici le cœur de ma position. S’interdire le savoir, c’est s’interdire d’argumenter depuis l’intérieur de la science.
Pour soutenir de façon convaincante qu’on peut sortir du modèle animal, il faut savoir précisément ce qu’il a donné, et ce que les alternatives peuvent remplacer. Refuser de regarder les études animales, c’est laisser le terrain à ceux qui défendent la vivisection. On se condamne à protester de l’extérieur, depuis la seule indignation morale, légitime, mais facile à disqualifier.
Et il y a mieux. La critique scientifique du modèle animal rejoint sa critique éthique. La transposition d’une espèce à l’autre échoue bien plus souvent qu’on ne le dit : quantité de cibles thérapeutiques « validées » chez la souris s’effondrent en clinique humaine. C’est précisément l’argument que porte un comité comme Antidote Europe fondé en 2004 par d’anciens chercheurs du CNRS, présidé scientifiquement et non militant au sens sentimental du terme : leur thèse n’est pas seulement « c’est cruel », c’est « aucune espèce n’est fiable pour prédire une autre espèce ». Ils proposent et financent des méthodes pertinentes pour l’humain : toxicogénomique sur cellules humaines, modèles in vitro, approches in silico, organoïdes et systèmes microphysiologiques (les fameux organ-on-chip). D’autres structures portent le même argumentaire scientifique ailleurs en Europe et aux États-Unis (Doctors Against Animal Experiments en Allemagne, le PCRM aux USA).
Autrement dit : être rigoureux sur les limites des données animales est à la fois de la bonne science et cohérent avec mon refus. L’humilité épistémique n’est pas une concession à l’adversaire, c’est mon meilleur outil. Et dans mon champ, la phyto-aromathérapie, où tant de mécanismes invoqués reposent sur de l’animal ou de l’in vitro extrapolés un peu vite à l’humain, cette vigilance est doublement opérante.
Un précédent qui éclaire : les cellules HeLa
Quand on cherche un modèle de résolution, l’analogie qui revient souvent, les données issues d’expériences criminelles, est trop disproportionnée pour être utile : elle écrase tout sous l’horreur et brouille même la validité scientifique.
Je préfère le cas des cellules HeLa : prélevées sans consentement sur Henrietta Lacks, devenues l’outil le plus universel de la biologie. La réponse de la communauté n’a pas été « cessons d’utiliser HeLa ». Elle a été : reconnaître l’origine, honorer la source, redistribuer une part des bénéfices, et surtout changer les normes de consentement en amont. Réparateur et tourné vers l’avenir, pas abstentionniste. C’est exactement la posture qui me semble juste.
Où je range, finalement, mon malaise
Je ne cherche pas à faire disparaître la gêne. Au contraire : une aisance totale à mobiliser ces données serait pire. Cette gêne, je la déplace. Elle ne dit pas « tu n’as pas le droit de citer ». Elle dit « cite en conscience, et pousse pour la suite ».
Concrètement, voilà ma règle de conduite :
privilégier la donnée humaine, clinique ou in vitro, quand elle suffit ;
mobiliser l’étude animale seulement quand c’est la meilleure preuve disponible et alors signaler ses limites, y compris translationnelles, pas seulement éthiques ;
donner visiblement leur place aux méthodes substitutives et aux structures qui les développent ;
citer en connaissance de cause, jamais machinalement.
Présenter ces études ne fait pas de moi un soutien à l’expérimentation animale. Refuser de les regarder ne sauve aucun animal, le tort est déjà fait, et affaiblit la cause que je porte. La pureté de provenance est un luxe que presque aucun savoir n’offre. La responsabilité, elle, se joue ailleurs : dans l’usage que je fais de ce savoir, et dans ce que je choisis de faire advenir à la place.
Pour une troisième compréhension du geste aromathérapeutique
1. Introduction
Le discours public sur la diffusion des huiles essentielles oscille presque toujours entre deux registres. Le premier est thérapeutique : on diffuse pour assainir l’air, dégager les voies respiratoires, faciliter le sommeil, agir sur l’humeur. Ce registre revendique des effets pharmacologiques précis, parfois étayés par des études cliniques de qualité variable, parfois extrapolés depuis des effets in vitro à des doses sans rapport avec l’exposition ambiante. Le second registre est cosmétique : on diffuse pour le plaisir, pour parfumer un intérieur, pour créer une ambiance. Ce registre ne revendique rien d’autre qu’une expérience sensorielle agréable.
Ces deux cadres ne sont pas faux. Ils captent chacun une dimension réelle du geste. Mais leur dualité, en occupant tout l’espace discursif, masque une troisième dimension qui me semble tout aussi intéressante que les deux premières, et plus rigoureuse, paradoxalement, parce qu’elle revendique moins.
L’hypothèse défendue dans cet article est la suivante : la diffusion d’huiles essentielles, dans son régime ambiant et à doses modérées, peut être pensée comme la réintroduction partielle d’un signal environnemental que la condition urbaine moderne a effacé. Ce cadre conceptuel diffère du cadre thérapeutique en ce qu’il ne prétend pas à un effet pharmacologique ciblé sur une pathologie ; il diffère du cadre cosmétique en ce qu’il enracine la pratique dans une physiologie évolutive identifiable. Il s’agit d’un cadre intermédiaire, plus modeste sur l’effet, plus précis sur le mécanisme.
Je propose d’examiner cette hypothèse en six étapes. La section 2 introduit l’énigme empirique du « bain de forêt » comme point d’ancrage. La section 3 examine puis écarte une analogie séduisante mais structurellement fausse, celle de la vitamine C, pour montrer pourquoi le cadre conceptuel correct n’est pas celui d’un nutriment caché. La section 4 reformule le cadre dans les termes de la xénohormèse et de l’exposome. La section 5 expose la spécificité neuroanatomique du canal olfactif, qui rend ce signal particulier parmi les autres signaux environnementaux possibles. La section 6 mobilise les données empiriques de la psychologie environnementale pour montrer que la « mémoire biophile » n’est pas une spéculation mais un phénomène mesurable. La section 7 examine les implications pour la pratique, ainsi que les limites et risques d’une telle reformulation.
2. Le bain de forêt comme énigme empirique
Le point de départ contemporain de cette réflexion est l’ensemble des travaux sur le shinrin-yoku (bain de forêt) menés depuis les années 2000 au Japon, principalement par Qing Li et son équipe à la Nippon Medical School. Dans une série d’études, Li a montré que des sujets exposés pendant plusieurs jours à un environnement forestier, typiquement deux à trois jours de marche en forêt, présentent une élévation mesurable et durable de l’activité de leurs cellules NK (natural killer), un type de lymphocytes impliqués dans la défense antivirale et antitumorale précoce (Li, 2010 ; Li et al., 2008).
Les chiffres rapportés varient selon les protocoles, mais l’ordre de grandeur typique se situe autour d’une augmentation de 30 à 50% de l’activité cytotoxique NK, avec une persistance mesurable jusqu’à un mois après l’exposition. Plusieurs hypothèses confondantes ont été testées et écartées : l’effet n’est pas attribuable à la qualité de l’air seule, ni à l’exercice physique, ni à la déconnexion numérique, puisque des groupes témoins exposés à des conditions équivalentes en milieu urbain n’ont pas présenté la même réponse.
L’hypothèse explicative qui s’est imposée par défaut est celle des phytoncides — terme japonais désignant l’ensemble des composés organiques volatils émis par les plantes, et notamment les monoterpènes comme l’α-pinène, le β-pinène, le limonène, et le 3-carène. L’idée est que l’inhalation chronique de ces molécules dans l’environnement forestier exerce une modulation immune, possiblement via des effets anti-inflammatoires et anti-stress (cortisol, axe HPA).
Cette explication reste partielle. L’attribution causale précise à une ou plusieurs molécules est difficile, parce que la matrice forestière comprend simultanément un cocktail moléculaire complexe, une stimulation multisensorielle (lumière diffractée, sons, humidité, microbiote aéroporté), et une réduction du stress urbain. Néanmoins, des études complémentaires sur l’inhalation contrôlée de phytoncides extraits ont montré des effets immuns mesurables (Li et al., 2009), ce qui rend l’hypothèse de la composante moléculaire vraisemblable sans en faire la seule explication.
Pour notre propos, ce qui importe n’est pas de trancher la question causale en détail, mais de constater qu’il existe un faisceau d’observations empiriques cohérentes selon lesquelles l’exposition à un environnement biotique riche produit chez l’humain des effets physiologiques mesurables et reproductibles, qu’on ne retrouve pas dans des environnements abiotiques équivalents.
3. Pourquoi l’analogie avec la vitamine C ne tient pas
Une intuition spontanée surgit face à ces données : si notre corps réagit favorablement à une exposition aux molécules végétales, n’est-ce pas parce qu’il en a besoin ? Et plus précisément : ne sommes-nous pas dans une configuration analogue à celle de la vitamine C, c’est-à-dire dépendants d’une molécule que nous ne synthétisons pas mais que notre alimentation ou notre environnement ancestral fournissait en permanence ?
Cette analogie est intuitivement séduisante, mais elle ne résiste pas à un examen mécanistique. La compréhension de ses limites est instructive parce qu’elle conduit à une reformulation plus juste du cadre conceptuel.
La vitamine C présente un ensemble de caractéristiques qui en font un nutriment essentiel au sens strict. Premièrement, l’espèce humaine a perdu la capacité de la synthétiser : le gène GULO (L-gulonolactone oxidase), qui catalyse la dernière étape de la biosynthèse de l’ascorbate, a été pseudogénisé chez nos ancêtres primates il y a environ 60 à 75 millions d’années (Drouin et al., 2011 ; Nishikimi & Yagi, 1991). Cette perte est documentée moléculairement et reproduite chez d’autres lignées (cobayes, certaines chauves-souris). Deuxièmement, l’organisme humain dispose d’une machinerie dédiée à la conservation de l’ascorbate : transporteurs spécifiques (SVCT1 et SVCT2), réabsorption rénale active, et intégration comme cofacteur dans plusieurs enzymes essentielles (hydroxylases du collagène, dopamine-β-hydroxylase, carnitine biosynthesis). Troisièmement, son absence prolongée produit un syndrome clinique reproductible — le scorbut — avec une histoire pathologique bien documentée.
L’α-pinène et les autres monoterpènes ne cochent aucune de ces cases. Premièrement, l’espèce humaine n’a jamais synthétisé ces molécules : les mammifères ne possèdent pas les terpène-synthases nécessaires, et il n’y a donc pas de « perte » de biosynthèse à compenser. Deuxièmement, l’organisme humain traite ces molécules comme des xénobiotiques classiques : oxydation par les cytochromes P450 (notamment CYP2B6) en métabolites comme le verbénol, la verbénone, et autres dérivés oxygénés ; conjugaison de phase II ; excrétion biliaire ou rénale (Salehi et al., 2019). Cette voie de clairance est l’inverse d’une voie de rétention. Troisièmement, aucun syndrome de carence n’a été identifié chez les populations historiquement éloignées des environnements riches en monoterpènes.
L’analogie avec la vitamine C est donc structurellement fausse. L’α-pinène n’est pas un nutriment caché. Et ce constat semble à première vue invalider l’idée d’un « besoin » physiologique de l’organisme pour ces molécules.
Mais c’est précisément la conclusion erronée à tirer. L’absence d’un statut de nutriment essentiel n’implique pas l’absence de pertinence physiologique. Elle invite à reformuler le cadre dans des termes différents.
4. Reformulation : signal environnemental et exposome
Le cadre conceptuel correct n’est pas celui du nutriment essentiel, mais celui du signal environnemental. Cette distinction est cruciale et permet d’articuler les données empiriques sans surenchérir sur leurs implications.
Un nutriment essentiel est une molécule dont l’absence produit une pathologie identifiable et dont la présence est intégrée à des voies métaboliques précises. Un signal environnemental, en revanche, est une molécule (ou une classe de molécules) dont la présence n’est pas requise pour le fonctionnement de base de l’organisme, mais qui module des paramètres physiologiques de manière biologiquement adaptative dans un environnement où elle est statistiquement attendue.
L’hypothèse de la xénohormèse, formulée par Howitz et Sinclair (2008), constitue le cadre théorique pertinent. Elle propose que les animaux ont co-évolué avec les plantes dans des environnements où les composés volatils végétaux signalent l’état de l’écosystème, et que les organismes hétérotrophes ont développé des mécanismes de réponse à ces signaux. Réponses qui peuvent inclure des effets adaptatifs (modulation immune, anti-inflammatoire, antioxydante) sans que la molécule en question soit pour autant un nutriment au sens strict. Les polyphénols (resvératrol, curcuminoïdes) et les monoterpènes entreraient dans cette catégorie de signaux environnementaux modulateurs.
Ce cadre est cohérent avec le concept plus large d’exposome introduit par Wild (2005) : l’ensemble des expositions environnementales auxquelles un organisme est soumis au cours de sa vie, qui complète l’information génétique pour expliquer les phénotypes de santé. L’exposome inclut la lumière, les microbes, les composés chimiques de l’air et de l’alimentation, et ce que je propose d’appeler à la suite de différents auteurs l’exposome moléculaire végétal — l’ensemble des composés volatils émis par les plantes auxquels un organisme est exposé.
Cette reformulation a une conséquence importante. Elle permet de prendre au sérieux l’hypothèse selon laquelle la condition urbaine moderne, en réduisant drastiquement l’exposition aux composés végétaux volatils, constitue un appauvrissement de l’exposome dont les effets sont nécessairement diffus, non spécifiques, et difficiles à isoler dans des études cliniques classiques — mais empiriquement plausibles. Les effets ne se manifesteront pas comme une « carence en α-pinène », mais comme une dérégulation cumulative de paramètres immuns, inflammatoires, et neurovégétatifs, qui interagit avec d’autres dimensions de l’exposome appauvri (microbiote, lumière, mouvement).
Dans ce cadre, la diffusion d’huiles essentielles peut être pensée non comme un soin ciblé ni comme un simple plaisir olfactif, mais comme une réintroduction partielle d’un signal environnemental attendu par notre physiologie évolutive. Cette formulation a l’avantage d’être épistémiquement modeste, elle ne revendique pas un effet curatif spécifique et empiriquement défendable, parce qu’elle s’appuie sur des phénomènes mesurables et non sur une analogie trompeuse.
5. La spécificité neuroanatomique du canal olfactif
Si l’on accepte le cadre du signal environnemental, une question subsiste : pourquoi la voie olfactive serait-elle particulièrement pertinente parmi toutes les voies sensorielles possibles de réintroduction d’un signal naturel ? La réponse tient à une particularité neuroanatomique de l’olfaction qui mérite d’être détaillée.
Les modalités sensorielles classiques (vision, audition, somesthésie, gustation) partagent une architecture commune : l’information sensorielle est relayée par le thalamus avant d’atteindre les aires corticales primaires, où elle est traitée puis intégrée à des aires associatives plus élaborées. Cette voie thalamique constitue un point de passage obligé qui structure la perception consciente.
L’olfaction, seule parmi les sens, ne suit pas cette architecture. L’information olfactive captée par l’épithélium olfactif est transmise au bulbe olfactif, qui projette directement sur le cortex piriforme, l’amygdale, le cortex entorhinal et certaines régions de l’hippocampe c’est-à-dire vers des structures limbiques impliquées dans la mémoire affective, l’évaluation hédonique, et la régulation du système nerveux autonome (Gottfried & Zald, 2005 ; Sela & Sobel, 2010). La voie thalamo-corticale existe, mais elle est subsidiaire et tardive dans le traitement.
Cette architecture explique plusieurs propriétés bien documentées de l’olfaction humaine : la difficulté à verbaliser les odeurs, la robustesse de la mémoire olfactive (souvent plus durable que les mémoires visuelles équivalentes), et surtout la capacité des stimuli olfactifs à déclencher des réponses émotionnelles et autonomes avant même que l’identification consciente ne soit achevée. Les phénomènes proustiens de réminiscence affective déclenchée par une odeur ne sont pas des curiosités littéraires ; ils sont des manifestations directes de cette architecture neuroanatomique particulière.
Pour notre propos, cette particularité a une conséquence importante : un stimulus olfactif est une voie privilégiée pour atteindre les structures qui modulent l’état physiologique interne, sans passer par l’analyse consciente. Là où une représentation visuelle de la nature (photographie, vidéo, fenêtre sur jardin) doit être décodée et identifiée pour produire ses effets, un stimulus olfactif agit plus directement.
Il faut ajouter une distinction de nature sémiotique qui me semble pertinente. Une représentation visuelle de la nature est, au sens peircien, un signe iconique ou symbolique : elle évoque la nature par ressemblance ou par convention, mais n’est pas elle-même la nature. Un stimulus olfactif issu d’une plante (que ce soit en forêt ou via une diffusion d’huile essentielle) est un signe indiciel : la molécule présente dans l’air est physiquement et chimiquement identique à celle émise par la plante, et il existe un lien causal réel entre le signe et ce qu’il signifie. Le cerveau qui reçoit le signal olfactif ne reçoit pas une représentation de la nature ; il reçoit un fragment chimique authentique de cette nature.
Cette précision sémiotique n’est pas une coquetterie philosophique : elle explique pourquoi le canal olfactif fait ce que les canaux purement représentationnels ne font pas, et pourquoi l’effet d’une diffusion ne peut pas être réduit à une « simulation » de la nature.
6. Données empiriques sur la latence biophile
L’hypothèse selon laquelle l’organisme humain conserve une réceptivité aux signaux environnementaux naturels, ce qu’on peut appeler à la suite de E.O. Wilson une biophilie (Wilson, 1984 ; Kellert & Wilson, 1993) n’est pas seulement une spéculation théorique. Plusieurs lignes de recherche empirique convergent vers un même constat : il existe chez l’humain contemporain une préférence pour les stimuli naturels qui se manifeste par des effets physiologiques et comportementaux mesurables, indépendamment de la déclaration consciente des sujets.
L’étude fondatrice est celle d’Ulrich (1984), publiée dans Science. Sur une cohorte de 46 patients ayant subi une cholécystectomie dans un hôpital de Pennsylvanie entre 1972 et 1981, Ulrich a comparé la récupération post-opératoire selon l’orientation de la chambre. Les patients dont la fenêtre donnait sur un bouquet d’arbres ont présenté une durée d’hospitalisation significativement plus courte (en moyenne 7,96 jours contre 8,71 jours, soit environ 0,75 jour de moins), une consommation moindre d’antalgiques puissants, et moins de complications post-opératoires mineures, comparés aux patients dont la fenêtre donnait sur un mur de briques. L’échantillon est limité, mais le protocole appariait soigneusement les variables confondantes (équipe soignante, étage, période, etc.), ce qui rend le résultat difficile à attribuer à autre chose qu’à la différence de vue.
Cette étude pionnière a été suivie par un corpus important de recherches en psychologie environnementale, dont les principales lignes peuvent être résumées ainsi.
Kaplan et Kaplan (1989) ont théorisé l’Attention Restoration Theory, selon laquelle les environnements naturels favorisent la récupération de l’attention dirigée (épuisée par les exigences cognitives urbaines) en sollicitant une forme d’attention « sans effort » (soft fascination). Plusieurs études expérimentales ont depuis montré que des promenades en environnement naturel améliorent les performances cognitives à des tâches d’attention dirigée, comparées à des promenades urbaines de durée et d’intensité physique équivalentes (Berman, Jonides & Kaplan, 2008).
Kuo et Sullivan (2001a, 2001b), dans une série d’études sur des résidents d’immeubles sociaux à Chicago, ont mis en évidence des corrélations entre la quantité de végétation visible depuis le logement et plusieurs indicateurs de santé mentale et de comportement social : moindre fatigue mentale, moindre agressivité interpersonnelle, et corrélation plus surprenante, moindre incidence de violences criminelles dans les zones plus végétalisées. Les protocoles statistiques contrôlent partiellement les variables socio-économiques confondantes.
Au-delà de ces études classiques, il existe également des observations comportementales fines, notamment en urbanisme et en éthologie urbaine, montrant que les piétons en ville présentent des préférences spatiales révélées pour les itinéraires comportant de la végétation, et qu’ils dévient leur trajectoire pour passer à proximité d’arbres, sans en avoir conscience explicite. Ces préférences révélées constituent, au sens où Daniel Kahneman et l’économie comportementale l’entendent, des données plus robustes que les préférences déclarées, parce qu’elles ne sont pas filtrées par la rationalisation post-hoc.
L’ensemble de ces données ne démontre pas mécanistiquement le lien causal entre exposition aux composés volatils végétaux et effets biophiles, ce sont des observations multimodales qui combinent vue, odeur, son, et association globale d’un environnement biotique. Mais elles établissent le fait empirique d’une réceptivité humaine persistante aux signaux naturels, qui opère en grande partie sous le seuil de la conscience délibérative.
7. Implications, limites et risques
Si l’on accepte l’ensemble du cadre développé jusqu’ici, la pratique de la diffusion d’huiles essentielles en intérieur urbain peut être pensée comme une réintroduction modeste d’un fragment de l’exposome moléculaire végétal absent de l’environnement urbain typique. Cette formulation a plusieurs avantages.
Elle évite la surenchère pharmacologique caractéristique d’une partie de la littérature aromathérapeutique, qui extrapole des effets in vitro à doses élevées vers des conditions ambiantes où les concentrations atteintes sont inférieures de plusieurs ordres de grandeur. À titre d’ordre de grandeur, l’air d’une forêt de conifères contient typiquement entre 0,5 et 5 μg/m³ d’α-pinène ; une diffusion atmosphérique modérée en intérieur peut atteindre des concentrations de l’ordre de 50 à 500 μg/m³, mais sur des durées limitées. Ces doses ne reproduisent pas les effets pharmacologiques systémiques étudiés en laboratoire à des concentrations de l’ordre du mg/kg, mais elles peuvent suffire à délivrer un signal modulateur perceptible, par les voies olfactives notamment.
Elle articule la pratique individuelle à une problématique collective. Si l’exposome moléculaire végétal urbain est appauvri, alors les solutions ne se limitent pas aux pratiques individuelles de diffusion : elles impliquent aussi la végétalisation urbaine, la perméabilité des espaces verts, l’aération des logements, la présence de plantes vivantes. La diffusion individuelle s’inscrit comme une réponse partielle parmi d’autres, et non comme un substitut à un changement environnemental plus large.
Plusieurs limites doivent cependant être posées clairement.
Premièrement, le cadre proposé est conceptuellement cohérent et empiriquement plausible, mais il n’est pas démonté à un niveau mécanistique fin. L’attribution causale précise des effets observés en forest bathing aux molécules volatiles, plutôt qu’à d’autres composantes de l’exposition, reste partielle. Les transpositions à des conditions de diffusion en intérieur sont encore moins étudiées spécifiquement.
Deuxièmement, il existe un risque de dérive interprétative qui doit être identifié. La pratique de la diffusion peut fonctionner de deux manières fonctionnellement distinctes : soit comme un rappel qui réactive la conscience d’un lien à la nature et qui s’articule à d’autres pratiques (sortie en forêt, jardinage, attention écologique) ; soit comme un palliatif qui rend la déconnexion urbaine plus supportable et tend à neutraliser la motivation à modifier l’environnement structurel. Les deux fonctions ne sont pas mutuellement exclusives et coexistent vraisemblablement selon les usages. Mais il importe que le praticien et le public soient conscients de cette ambiguïté fonctionnelle, et que la pratique soit cadrée pour favoriser la première fonction plutôt que la seconde.
Troisièmement, le cadre développé concerne spécifiquement la diffusion ambiante à doses modérées. Il ne s’applique pas tel quel aux régimes pharmacologiques classiques de l’aromathérapie (voie orale, application cutanée concentrée, suppositoires, diffusion intensive ciblée), qui relèvent d’une logique différente, pharmacologie standard avec courbes dose-réponse, indications cliniques précises, contre-indications, métabolisme hépatique sollicité. Confondre les deux régimes est un piège fréquent : justifier le second par la rationalité du premier, ou disqualifier le premier sur la base des limites du second, sont deux erreurs symétriques.
8. Conclusion
L’argument central de cet article tient en une formule : la diffusion d’huiles essentielles, dans son régime ambiant, ne reconstitue pas la nature absente, elle réactive la mémoire qu’on en a. Cette mémoire, dont les recherches en psychologie environnementale montrent qu’elle est intacte sous l’urbain et opère en partie sous le seuil de la délibération, n’est pas une essence mystique ; c’est l’ensemble des dispositifs neurophysiologiques calibrés par plusieurs centaines de milliers d’années de coévolution avec un environnement biotique riche.
Cette reformulation a une valeur épistémique : elle remplace une revendication d’effet pharmacologique faiblement étayée par une revendication de modulation environnementale mieux fondée. Elle a aussi une valeur pratique : elle articule le geste individuel à une question collective (l’aménagement urbain), et elle permet de cadrer la pratique pour qu’elle fonctionne comme un rappel et non comme un palliatif.
Elle a enfin une valeur réflexive pour la pratique aromathérapeutique elle-même : en distinguant clairement le régime pharmacologique du régime environnemental de la diffusion, elle permet de défendre chacun sur ses propres bases sans que les limites de l’un fragilisent l’autre. Cette distinction conceptuelle, banale en pharmacologie mais souvent absente dans le discours aromathérapeutique courant, me semble une condition de la rigueur épistémique de la discipline.
Références bibliographiques
Berman, M. G., Jonides, J., & Kaplan, S. (2008). The cognitive benefits of interacting with nature. Psychological Science, 19(12), 1207–1212.
Drouin, G., Godin, J. R., & Pagé, B. (2011). The genetics of vitamin C loss in vertebrates. Current Genomics, 12(5), 371–378.
Gottfried, J. A., & Zald, D. H. (2005). On the scent of human olfactory orbitofrontal cortex: meta-analysis and comparison to non-human primates. Brain Research Reviews, 50(2), 287–304.
Howitz, K. T., & Sinclair, D. A. (2008). Xenohormesis: sensing the chemical cues of other species. Cell, 133(3), 387–391.
Kahneman, D. (2011). Thinking, Fast and Slow. Farrar, Straus and Giroux.
Kaplan, R., & Kaplan, S. (1989). The Experience of Nature: A Psychological Perspective. Cambridge University Press.
Kaplan, S. (1995). The restorative benefits of nature: Toward an integrative framework. Journal of Environmental Psychology, 15(3), 169–182.
Kellert, S. R., & Wilson, E. O. (Eds.). (1993). The Biophilia Hypothesis. Island Press.
Kuo, F. E., & Sullivan, W. C. (2001a). Aggression and violence in the inner city: Effects of environment via mental fatigue. Environment and Behavior, 33(4), 543–571.
Kuo, F. E., & Sullivan, W. C. (2001b). Environment and crime in the inner city: Does vegetation reduce crime? Environment and Behavior, 33(3), 343–367.
Li, Q. (2010). Effect of forest bathing trips on human immune function. Environmental Health and Preventive Medicine, 15(1), 9–17.
Li, Q., Morimoto, K., Kobayashi, M., Inagaki, H., Katsumata, M., Hirata, Y., Hirata, K., Suzuki, H., Li, Y. J., Wakayama, Y., Kawada, T., Park, B. J., Ohira, T., Matsui, N., Kagawa, T., Miyazaki, Y., & Krensky, A. M. (2008). Visiting a forest, but not a city, increases human natural killer activity and expression of anti-cancer proteins. International Journal of Immunopathology and Pharmacology, 21(1), 117–127.
Li, Q., Nakadai, A., Matsushima, H., Miyazaki, Y., Krensky, A. M., Kawada, T., & Morimoto, K. (2009). Phytoncides (wood essential oils) induce human natural killer cell activity. Immunopharmacology and Immunotoxicology, 28(2), 319–333.
Morin, E. (1977–2004). La Méthode (6 volumes). Éditions du Seuil.
Nishikimi, M., & Yagi, K. (1991). Molecular basis for the deficiency in humans of gulonolactone oxidase, a key enzyme for ascorbic acid biosynthesis. American Journal of Clinical Nutrition, 54(6 Suppl), 1203S–1208S.
Salehi, B., Upadhyay, S., Erdogan Orhan, I., Kumar Jugran, A., L. D. Jayaweera, S., A. Dias, D., Sharopov, F., Taheri, Y., Martins, N., Baghalpour, N., Cho, W. C., & Sharifi-Rad, J. (2019). Therapeutic potential of α- and β-pinene: A miracle gift of nature. Biomolecules, 9(11), 738.
Sela, L., & Sobel, N. (2010). Human olfaction: a constant state of change-blindness. Experimental Brain Research, 205(1), 13–29.
Ulrich, R. S. (1984). View through a window may influence recovery from surgery. Science, 224(4647), 420–421.
Wild, C. P. (2005). Complementing the genome with an « exposome »: The outstanding challenge of environmental exposure measurement in molecular epidemiology. Cancer Epidemiology, Biomarkers & Prevention, 14(8), 1847–1850.
Wilson, E. O. (1984). Biophilia. Harvard University Press.
Dans le domaine de la pharmacognosie clinique, le concept de totum désigne l’intégralité des molécules aromatiques d’une plante. Par extension, en aromathérapie la notion de totum recouvre simplement le fait pour une huile essentielle de contenir les principes actifs volatils obtenus par la distillation à la vapeur d’eau. Donc juste une partie infime du totum de la plante, ce qui en soi s’éloigne déjà de la définition originelle. Parler du totum en aromathérapie est donc déjà un non-sens même si une huile essentielle peut contenir de quelques dizaines à plusieurs centaines de constituants.
L’huile essentielle de tea tree (Melaleuca alternifolia) constitue un exemple intéressant pour parler de la notion de totum. Si le paradigme traditionnel de l’aromathérapie soutient que la synergie naturelle du totum de l’huile essentielle est systématiquement supérieure à l’action d’un principe actif isolé, la recherche contemporaine impose une vision plus nuancée.
L’efficacité du mélange dépend strictement du contexte thérapeutique et du pathogène ciblé, révélant que le totum n’est pas une réponse universelle mais un équilibre dynamique parfois perfectible.
L’huile de Melaleuca alternifolia se distingue par une composition riche d’environ 100 composés, dominée par plusieurs familles biochimiques majeures. Le terpinène-4-ol est le constituant prédominant (environ 40 %), classé parmi les monoterpénols.
On retrouve également dans ce totum :
D’autres monoterpénols : l’alpha-terpinéol.
Des oxydes (éthers) : le 1,8-cinéole (eucalyptol).
Des monoterpènes : l’alpha-terpinène, le gamma-terpinène, le sabinène et le terpinolène.
Si le terpinène-4-ol est souvent désigné comme l’agent actif principal, son interaction avec les molécules minoritaires définit la capacité biocide réelle de l’huile.
Le tea-tree face au Demodex
Le parasite Demodex (D. folliculorum), un acarien ectoparasite colonisant les follicules pileux et les glandes sébacées humaines, est souvent impliqué dans les blépharites et la rosacée. Dans ce contexte précis, l’approche par le totum se heurte à des limites pharmacologiques. L’étude de Tighe et al. (2013) a démontré que le terpinène-4-ol isolé est plus efficace pour éradiquer les acariens que l’huile essentielle entière.
Effet antagoniste avéré : L’interaction entre le terpinène-4-ol (le principal agent acaricide de l’huile d’arbre à thé) et l’alpha-terpinéol crée une « synergie négative » ou un effet antagoniste dans le contexte de l’éradication des acariens Demodex.
Augmentation du temps de survie du parasite : Lorsque le terpinène-4-ol est mélangé à l’alpha-terpinéol, l’efficacité acaricide est inhibée et le temps de survie des acariens augmente de façon significative comparativement à l’utilisation du terpinène-4-ol de manière isolée. Par exemple, lors de tests in vitro, la combinaison de ces deux composants a donné un temps de survie doublé du parasite, démontrant ainsi que l’alpha-terpinéol réduit l’efficacité du terpinène-4-ol.
Explication biochimique et physique : Cet antagonisme pourrait provenir du fait que la présence d’alpha-terpinéol modifie la solubilité ou la dynamique de pénétration du mélange. Cette interférence empêcherait le terpinène-4-ol de pénétrer efficacement la cuticule de l’acarien ou son système respiratoire.
Supériorité de l’utilisation isolée : Les chercheurs concluent que l’utilisation du terpinène-4-ol seul est supérieure car elle élimine les effets indésirables et antagonistes d’autres ingrédients, maximisant ainsi sa puissance brute pour détruire les Demodex.
Ces effets antagonistes ne sont malheureusement pas compensés par la synergie positive que procure l’association du terpinolène-4-ol et du terpinolène un autre principe actif du tea-tree.
On a donc pour le tea-tree face au Demodex deux effets opposés
Une coopération bénéfique entre le terpinène-4-ol et le terpinolène, renforçant l’action acaricide.
Un effet pénalisant de l’alpha-terpinéol (pourtant puissant lorsqu’il est seul) qui réduit l’efficacité du terpinène-4-ol contre le parasite lorsqu’ils sont combinés.
Pour le traitement de la démodécie, l’utilisation d’un extrait enrichi ou d’un isolat de terpinène-4-ol est donc préférable. Cette « aromathérapie de précision » permet de s’affranchir des interactions antagonistes naturelles pour maximiser la puissance thérapeutique.
La force de la synergie contre les bactéries et les virus
À l’inverse, face aux bactéries et aux virus, la complexité du totum redevient son principal atout. L’activité antibactérienne majeure contre Staphylococcus aureus (notamment le SARM, résistant à la méticilline) et Escherichia coli. Pour le SARM, les valeurs de concentration minimale inhibitrice (MIC) et de concentration minimale bactéricide (MBC) sont particulièrement basses, se situant respectivement entre 0,25 % et 0,5 %.
Le mécanisme d’action est multidimensionnel grâce à la complémentarité des monoterpénols et des oxydes :
Perméabilisation membranaire : Les molécules lipophiles perturbent l’intégrité de la membrane plasmique.
Fuites ioniques : Cette altération provoque une perte critique de potassium.
Inhibition respiratoire : Le mélange inhibe spécifiquement la respiration glucose-dépendante de la bactérie, entraînant sa mort.
Sur le plan antiviral, le tea tree se montre particulièrement efficace contre les virus enveloppés (Grippe, HSV-1). Les recherches (Garrozzo 2009, Koch 2007) indiquent que le terpinène-4-ol agit lors de la phase d’adsorption (attachement à la membrane cellulaire). En interrompant les protéines de surface comme l’hémagglutinine, le totum empêche l’entrée du virus dans la cellule hôte, une action plus complexe à obtenir avec une molécule isolée.
Synthèse des interactions
Le mélange de molécules aromatiques ne produit pas systématiquement une amélioration des propriétés. On décrit quatre états d’interaction biochimique :
Type d’interaction
Effet observé
Indifférence
L’activité est indépendante de la présence d’une autre molécule.
Addition
L’effet global est égal à la somme des effets individuels (cumulatif).
Synergie
L’effet est significativement supérieur à la somme des composants.
Antagonisme
La présence d’une molécule diminue l’activité d’une autre.
Le tea tree illustre parfaitement cette dualité. Il bénéficie d’une synergie puissante contre les bactéries, mais subit un antagonisme marqué face au Demodex. En pratique les phénomènes d’indifférence ou d’antagonisme sont souvent ignorés postulant à tort une synergie universelle.
Vers une aromathérapie de précision
Le totum n’est pas une « potion magique » mais un complexe biochimique dont l’efficacité est strictement corrélée à la cible biologique. Pour une meilleure pratique on doit adopter une démarche rigoureuse et honnête qui nous nous permet de choisir les principes actifs pertinents et se rappeler la question fondamentale :
Quels principes actifs on cherche à mobiliser, et quelle forme d’extraction permet d’y accéder ?
Références scientifiques
Étude sur le terpinène-4-ol et les acariens (Demodex) :
Tighe, S., Gao, Y.-Y., & Tseng, S. C. G. (2013). Terpinen-4-ol is the most active ingredient of tea tree oil to kill Demodex mites. Translational Vision Science & Technology, 2(7), 2. https://doi.org/10.1167/tvst.2.7.2
Études sur le terpinène-4-ol, les huiles essentielles et la phase d’adsorption virale :
Garozzo, A., Timpanaro, R., Bisignano, B., Furneri, P. M., Bisignano, G., & Castro, A. (2009). In vitro antiviral activity of Melaleuca alternifolia essential oil. Letters in Applied Microbiology, 49(6), 806–808. https://doi.org/10.1111/j.1472-765X.2009.02740.x
Koch, C., Reichling, J., Schneele, J., & Schnitzler, P. (2008). Inhibitory effect of essential oils against herpes simplex virus type 2. Phytomedicine, 15(1-2), 71–78. https://doi.org/10.1016/j.phymed.2007.09.003(Pré-publiée en 2007, décrivant l’action sur l’adsorption virale).
On imagine souvent qu’il est complexe, mais si on prend le temps de poser tranquillement les bases, il devient facilement compréhensible !
Le cycle ovarien révèle un processus biologique complexe dont la temporalité réelle dépasse largement la fenêtre conventionnelle de 28 jours. Bien que les courbes hormonales se focalisent sur cette période, l’ovule entame son voyage plus de trois mois avant l’ovulation. Le cycle est régi par une orchestration précise entre l’hypothalamus, l’hypophyse et les ovaires, passant par des phases de régulation négative et positive. Au-delà de la reproduction, les œstrogènes et la progestérone agissent comme des régulateurs systémiques globaux, protégeant les systèmes osseux, cardiovasculaires et neurologiques. La ménopause ne marque donc pas seulement la fin de la fertilité, mais la perte d’une protection multi-organique essentielle.
1. Au-delà des 28 Jours
Le cycle ovarien est l’aboutissement d’un processus de maturation folliculaire de longue durée. Il est crucial de distinguer le stock initial de la phase de maturation finale.
Le stock folliculaire et son évolution
Vie fœtale : Un pic de 7 millions de follicules primordiaux est atteint.
Naissance : Le stock tombe à environ 1 million.
Puberté : Il n’en reste que 400 000.
Processus d’atrésie : Avant la puberté, les follicules qui atteignent le stade antral meurent par manque de stimulation hormonale (FSH), expliquant la diminution constante du stock.
Anatomie du follicule
Un follicule est une structure protectrice et nourricière pour l’ovocyte, composée de deux types de cellules clés :
Cellules de la thèque : Situées à l’extérieur et vascularisées, elles reçoivent les nutriments et produisent des androgènes sous l’influence de la LH.
Cellules de la granulosa : Non vascularisées, elles nourrissent l’ovocyte, produisent l’AMH (Hormone Anti-Müllérienne) et convertissent les androgènes en œstrogènes sous l’influence de la FSH.
2. Mécanismes de maturation et régulation de l’AMH
Le développement folliculaire se déroule en plusieurs étapes, dont les premières sont indépendantes des hormones cycliques (FSH/LH).
Activation spontanée : Des follicules se « réveillent » de manière aléatoire du stock primordial vers les stades primaire et secondaire.
Rôle de l’AMH : Produite par les follicules en croissance, elle exerce un contrôle inhibiteur :
Elle maintient les autres follicules en état de dormance.
Elle ralentit la maturation des follicules déjà activés pour éviter un épuisement prématuré du stock.
Stade Antral : À ce stade, le follicule devient « hormono-dépendant ». Sans FSH, il subit une atrésie (mort cellulaire).
3. Dynamique hormonale du cycle mensuel
Le cycle est une alternance de rétroactions entre les hormones ovariennes et l’axe hypothalamus-hypophyse.
La phase folliculaire (régulation négative)
Production basale de LH : Indispensable dès le début pour permettre aux cellules de la thèque de produire les androgènes nécessaires.
Sécrétion de FSH : Stimule la maturation des follicules antraux.
Production d’œstrogènes : Les cellules de la granulosa transforment les androgènes en œstrogènes via l’aromatase.
Rétroaction négative : L’augmentation des œstrogènes signale au cerveau de réduire la production de FSH.
Sélection du Follicule Dominant : Seul le follicule le plus sensible à la FSH (activité aromatase intense) survit à la baisse de cette hormone ; les autres meurent.
L’Ovulation (Régulation positive)
Arrivé à un certain seuil d’œstrogènes, le cerveau change de comportement et passe en régulation positive. L’augmentation des œstrogènes provoque alors un pic de LH et un pic de FSH, déclenchant l’expulsion de l’ovule.
La phase lutéale
Le corps jaune : Après l’ovulation, les cellules de la granulosa se transforment en corps jaune.
Progestérone : Sous l’influence de la LH basale, le corps jaune produit de la progestérone.
Retour à la régulation négative : La combinaison œstrogènes + progestérone indique au cerveau que l’ovulation a eu lieu, abaissant la FSH et la LH.
Fin de Cycle : En l’absence de fécondation, le corps jaune s’atrophie. La chute des hormones lève l’inhibition sur la FSH, permettant le début d’un nouveau cycle.
4. Rôles systémiques des hormones ovariennes
Les hormones ne se limitent pas à la reproduction ; elles sont des régulateurs globaux de la santé féminine.
Système
Action des Œstrogènes
Action de la Progestérone
Reproduction
Prolifération de l’endomètre, fluidification de la glaire cervicale.
Transformation sécrétoire de l’endomètre, épaississement de la glaire (fermeture du col).
Système Osseux
Inhibition des ostéoclastes (prévention de l’ostéoporose).
N/A
Cardiovasculaire
Vasodilatation, effet anti-inflammatoire sur l’endothélium, profil lipidique protecteur.
N/A
Système Nerveux
Modulation de la sérotonine et dopamine (humeur, mémoire).
Effet sédatif et anxiolytique (action sur le système nerveux central).
Autres
Maintien de l’élasticité cutanée, récupération musculaire.
Élévation de la température (+0,5°C), immunomodulation (protection de l’embryon).
5. l’Équilibre hormonal
L’équilibre entre œstrogènes et progestérone est défini par une complémentarité séquentielle :
Les œstrogènes se situent dans le registre de l’activation et de la préparation.
La progestérone se situe dans le registre de la stabilisation et de la protection.
La compréhension de ces mécanismes souligne que les déséquilibres, tels que la chute brutale de progestérone, ont des répercussions directes sur le bien-être (syndrome prémenstruel, troubles de l’humeur) et que la transition vers la ménopause représente un défi physiologique majeur touchant l’ensemble de l’organisme.
L’observation est connue de longue date des praticien·ne·s expérimentés : les huiles essentielles issues de plantes poussant dans des milieux méditerranéens, semi-arides ou montagneux présentent en général des profils chimiques plus riches, plus complexes et des propriétés thérapeutiques plus marquées que celles issues de cultures intensives en milieu tempéré humide. Cette observation empirique souvent traduite par la notion floue de qualité de terroir a toutefois une base biologique précise et documentée.
Comprendre les mécanismes par lesquels le stress hydrique façonne la composition chimique des huiles essentielles, c’est se donner les outils pour raisonner différemment ses prescriptions en allant au-delà de la lecture d’une simple fiche technique, du simple nom botanique ou du chémotype.
1. La fermeture stomatique comme point de départ d’une cascade métabolique
En conditions de stress hydrique (sol sec, faible rétention en eau, déficit en vapeur d’eau atmosphérique) la plante déclenche une réponse conservatoire immédiate, la fermeture de ses stomates, ces pores foliaires par lesquels s’effectuent les échanges gazeux avec l’environnement. Cette fermeture limite les pertes en eau par transpiration, ce qui est vital. Mais elle a une conséquence métabolique profonde, souvent sous-estimée.
En réduisant les échanges gazeux, la fermeture stomatique coupe l’approvisionnement en CO₂ disponible pour la photosynthèse. La plante se retrouve dans une situation paradoxale, la lumière solaire continue de frapper ses feuilles, chargeant l’appareil photosynthétique en énergie sous forme de pouvoir réducteur (NADPH) et d’intermédiaires carbonés, mais cette énergie ne peut plus être orientée vers la croissance et la production de biomasse. Il y a un excès de pouvoir réducteur sans débouché dans le métabolisme primaire.
La cellule végétale doit impérativement évacuer ce surplus sous peine de dommages oxydatifs graves. La solution consiste donc à réorienter massivement le flux métabolique vers le métabolisme secondaire. C’est le mécanisme décrit sous le terme de Carbon Overflow Hypothesis. La limitation devient paradoxalement un moteur de richesse chimique.
2. Les deux usines à terpènes sous pression
Les terpènes, constituants majoritaires de la plupart des huiles essentielles, sont biosynthétisés via deux voies métaboliques compartimentées dans la cellule végétale.
La voie MEP (voie plastidiale)
Localisée dans les chloroplastes, la voie MEP (2-C-méthyl-D-érythritol 4-phosphate) produit principalement les monoterpènes (C10) et les diterpènes. Les monoterpènes (linalol, alpha-pinène, 1,8-cinéole, camphre, pour n’en citer que quelques-uns) représentent la fraction volatile dominante de la majorité des huiles essentielles. En conditions de stress hydrique, la voie MEP s’emballe, c’est elle qui absorbe prioritairement l’excès de NADPH et d’intermédiaires pyruvate/G3P accumulés par la photosynthèse contrainte.
La voie MVA (voie cytosolique)
Localisée dans le cytosol et le réticulum endoplasmique, la voie MVA (mévalonate) produit principalement les sesquiterpènes (C15) et les triterpènes. Moins directement sollicitée par l’excès de pouvoir réducteur chloroplastique, elle intervient dans un second temps.
Le crosstalk entre compartiments
Ce qui est particulièrement remarquable dans les situations de stress sévère, c’est que les intermédiaires biochimiques des deux voies normalement strictement séparées par les membranes chloroplastiques débordent d’un compartiment à l’autre. Ce crosstalk entre les voies MEP et MVA produit des molécules hybrides et allonge considérablement la diversité chimique de l’huile essentielle. Le résultat n’est pas seulement une production quantitativement plus importante, mais surtout une architecture chimique qualitativement différente, plus complexe, plus diversifiée, conçue pour répondre à de multiples agressions simultanées.
3. Pourquoi la plante les produit
Comprendre les rôles biologiques des terpènes produits sous stress éclaire directement leur potentiel thérapeutique. Ces molécules ne sont pas des sous-produits du métabolisme, elles ont des fonctions précises de protection cellulaire documentées.
Protection des membranes cellulaires
En conditions de déshydratation, les membranes plasmiques perdent leur fluidité et leur intégrité structurale. Certains terpènes volatils, notamment l’isoprène, le plus simple des terpènes, s’intercalent dans les bicouches lipidiques membranaires et maintiennent leur fluidité dans des conditions extrêmes de stress thermique et hydrique combinés. Ce mécanisme est analogue au rôle du cholestérol dans les membranes animales, un régulateur de fluidité membranaire. Il est documenté chez de nombreuses espèces aromatiques méditerranéennes.
Piégeage des radicaux libres
Le stress hydrique s’accompagne invariablement d’une augmentation de la production de ROS (Reactive Oxygen Species) dans les cellules végétales, en particulier dans les chloroplastes dont le fonctionnement est perturbé. Les monoterpènes et les phénylpropanoïdes aromatiques agissent comme piégeurs directs de ces radicaux libres, limitant les dommages oxydatifs sur les protéines, les lipides membranaires et l’ADN. Cette activité antioxydante endogène de la plante, conçue pour se protéger elle-même, est précisément ce que nous exploitons à des fins thérapeutiques lorsque nous prescrivons ces molécules.
Thermoprotection de l’appareil photosynthétique
Le stress hydrique et le stress thermique sont fréquemment cooccurrents dans les milieux méditerranéens et montagnards. Les terpènes volatils émis par les feuilles forment une couche gazeuse périfoliaire qui contribue à moduler leur température par effet de refroidissement évaporatif protégeant ainsi les complexes protéiques de l’appareil photosynthétique contre la dénaturation. Ce mécanisme, longtemps débattu, est aujourd’hui bien documenté chez les espèces du genre Quercus, Pinus et plusieurs Lamiacées aromatiques.
Ces trois fonctions, stabilisation membranaire, piégeage des ROS, thermoprotection, expliquent mécanistiquement pourquoi les huiles essentielles de milieux contraignants présentent des activités antioxydantes, anti-inflammatoires et protectrices souvent supérieures à celles de leurs équivalents cultivés en conditions optimisées. La plante a synthétisé ces molécules pour se défendre, nous en empruntons les outils.
4. La nuance entre stress modéré et stress sévère
Une erreur fréquente consiste à conclure que plus la plante est stressée, meilleure est l’huile essentielle. La réalité biologique est plus nuancée et impose une vigilance pratique importante.
Le stress modéré chronique : moteur de richesse
Un stress hydrique modéré et chronique, celui d’un sol rocailleux, drainant, à faible rétention en eau, dans un contexte méditerranéen ou montagnard, déclenche la réorientation métabolique décrite précédemment. La plante survit, s’adapte, et oriente durablement son métabolisme vers la production de métabolites secondaires complexes. C’est le profil des terroirs qui produisent des huiles essentielles de haute densité thérapeutique comme la lavande d’altitude en Provence, la sarriette du Verdon, le thym du Causse ou l’origan méditerranéen.
Le stress sévère aigu : dérèglements et risques
Un stress hydrique sévère et brutal, sécheresse exceptionnelle, épisode de canicule prolongée , produit des effets différents et moins favorables. Les études sur Rosmarinus officinalis en sont l’illustration. Un stress modéré augmente la teneur en 1,8-cinéole et en alpha-pinène, des molécules bien tolérées et aux propriétés documentées, tandis qu’un stress sévère favorise l’accumulation de camphre, une molécule chimiquement plus stable, moins coûteuse à synthétiser en situation d’urgence, mais potentiellement plus toxique aux doses usuelles et contre-indiquée dans plusieurs populations.
Cette observation a une implication pratique directe, une huile essentielle de romarin issue d’une région ayant subi une sécheresse exceptionnelle peut présenter une teneur en camphre significativement supérieure à la normale, avec des conséquences réelles sur son profil de sécurité. La lecture de l’analyse GC/MS du lot concerné devient alors non pas un détail administratif, mais une information cliniquement pertinente.
Le cas de Lavandula angustifolia
Chez la lavande vraie, la dynamique est différente et mérite d’être précisée. À partir de 800 à 1000 mètres d’altitude, la plante cumule plusieurs formes de stress simultanés, stress hydrique chronique sur sol drainant et rocailleux, fortes amplitudes thermiques jour/nuit, et exposition accrue aux ultraviolets. Cette combinaison met la voie MEP sous pression soutenue et prolongée.
L’excès d’énergie pousse la biosynthèse jusqu’au terme de la chaîne réactionnelle. Le linalol, produit en abondance, est massivement estérifié pour former de l’acétate de linalyle. Cette estérification est elle-même favorisée par les basses températures nocturnes fréquentes en altitude. Le profil résultant riche en acétate de linalyle, avec un rapport élevé acétate de linalyle/linalol confère à ces huiles essentielles leurs propriétés spasmolytiques, anxiolytiques et sédatives caractéristiques.
À l’inverse, dans une culture de plaine irriguée, la biosynthèse s’arrête plus tôt dans la chaîne, davantage de linalol brut, moins d’acétate de linalyle. Les deux molécules ont des propriétés pharmacologiques distinctes, le linalol étant plus anxiolytique et antibactérien, l’acétate de linalyle plus sédatif et antispasmodique et ces différences de ratio ont des implications thérapeutiques concrètes.
Cette nuance stress modéré/sévère invalide tout raisonnement simpliste du type « stress = qualité ». Elle plaide pour une lecture fine des conditions climatiques du terroir et pour une analyse lot par lot, surtout dans les années climatiquement atypiques.
5. La notion de terroir chimique
La notion de terroir est empruntée à la viticulture, où elle désigne l’ensemble des facteurs pédologiques, climatiques et topographiques qui conditionnent la composition et les caractéristiques organoleptiques d’un vin. Appliquée aux huiles essentielles, cette notion repose sur des mécanismes biologiques précis qui viennent d’être décrites.
Un terroir sec, rocailleux, à faible rétention en eau, soumet la plante à un stress hydrique chronique modéré qui stimule durablement la production de métabolites secondaires et oriente la composition chimique vers des profils spécifiques. Ce n’est pas le sol en lui-même qui confère les propriétés, mais l’ensemble des contraintes que ce sol impose à la plante et les réponses adaptatives que ces contraintes suscitent.
Une lavande de haute altitude en Provence, une sarriette du Verdon, un thym du Causse ou un origan de Corse ne présentent pas les mêmes profils chimiques que leurs équivalents cultivés en plaine irriguée et cette différence n’est pas un argument de vente, c’est le reflet mesurable d’histoires biologiques distinctes.
La variabilité inter-annuelle
La notion de terroir chimique ouvre également une réflexion sur la variabilité inter-annuelle d’une huile essentielle issue d’un même producteur, d’une même parcelle. Les conditions hydriques varient d’une année à l’autre. Une année à pluviométrie abondante produira des huiles essentielles chimiquement différentes d’une année de sécheresse modérée, qui elle-même sera différente d’une année de sécheresse exceptionnelle.
Cette variabilité est documentée et prévisible dans ses mécanismes. Elle plaide pour une pratique systématique et non optionnelle de lecture des analyses GC/MS lot par lot. Non pas pour vérifier une conformité administrative à une norme de composition, mais pour connaître réellement l’outil que l’on utilise avant de l’utiliser.
6. Implications pratiques pour la pratique
La provenance comme donnée clinique
La première implication est simple à formuler mais profonde dans ses conséquences. La provenance géographique et les conditions climatiques d’une huile essentielle ne sont pas des informations anecdotiques ou des éléments de contexte. Elles conditionnent directement la composition chimique et donc les propriétés thérapeutiques ainsi que le profil de sécurité via les mécanismes de réponse au stress hydrique que décrits ci-dessus.
Cela signifie que deux huiles essentielles portant le même nom botanique, le même chémotype, distillées de la même partie de plante, peuvent présenter des propriétés thérapeutiques significativement différentes si leurs conditions de culture diffèrent. Le nom botanique est nécessaire mais insuffisant.
La lecture GC/MS comme outil de raisonnement
L’analyse chromatographique d’un lot n’est pas seulement un outil de contrôle qualité ou de détection de fraude. C’est un outil de raisonnement clinique. Les ratios entre molécules (acétate de linalyle/linalol, 1,8-cinéole/alpha-pinène/camphre, thymol/carvacrol) sont des reflets directs des conditions de croissance et des réponses adaptatives de la plante. Savoir les lire, c’est pouvoir anticiper le comportement thérapeutique et le profil de sécurité du lot que l’on va utiliser.
Valoriser les milieux contraignants sans romantisme
Comprendre la biologie du stress hydrique justifie de valoriser les huiles essentielles issues de milieux naturels contraignants non pas par romantisme naturaliste ou par réflexe de « naturel = meilleur », mais parce que le stress hydrique chronique modéré est un moteur biologique de richesse chimique et de complexité moléculaire que les conditions de culture optimisées pour le rendement agricole ne reproduisent pas.
Le rendement à l’hectare est inversement corrélé à la densité thérapeutique dans ce contexte. La plante sous contrainte produit peu de biomasse mais beaucoup de métabolites secondaires par gramme de feuille. Le coût supérieur des huiles essentielles issues de ces milieux est le reflet direct de cette réalité économique et biologique.
Suivre la variabilité climatique
Pour les praticien·ne·s qui travaillent avec des producteurs identifiés et des millésimes, il est pertinent de suivre les conditions climatiques des années de récolte. Une année exceptionnellement sèche dans une région de production habituellement modérément stressée peut produire des huiles essentielles atypiques, plus chargées en certaines molécules qui méritent une attention particulière à la lecture de l’analyse et aux conditions d’utilisation.
Conclusion
La biologie du stress hydrique offre aux praticien·ne·s quelque chose de précieux, un cadre mécanistique pour raisonner la qualité des huiles essentielles au-delà des classifications habituelles. Elle transforme des observations empiriques, « la lavande d’altitude est plus fine », « les plantes méditerranéennes sont plus aromatiques » en faits biologiques explicables et prédictibles.
Elle invite surtout à concevoir la molécule non plus comme un simple principe actif isolé, mais comme l’expression chimique d’une histoire, celle d’une plante qui a dû s’adapter à un environnement précis, mobiliser ses ressources, produire ses outils de survie. Ce que nous prescrivons, c’est en partie cette capacité d’adaptation et comprendre comment elle se construit nous rend meilleur·e·s.
Références & approfondissements recommandés
Gershenzon J. (1994). Metabolic costs of terpenoid accumulation in higher plants. Journal of Chemical Ecology.
Loreto F. & Schnitzler J.P. (2010). Abiotic stresses and induced BVOCs. Trends in Plant Science.
Bakkali F. et al. (2008). Biological effects of essential oils – a review. Food and Chemical Toxicology.
Munné-Bosch S. & Alegre L. (2000). Changes in carotenoids, tocopherols and diterpenes during drought and recovery, and the biological significance of chlorophyll loss in Rosmarinus officinalis plants. Planta, 210(6), 925-931.
Sangwan N.S. et al. (2001). Regulation of essential oil production in plants: developmental and environmental factors as determinants. Plant Growth Regulation.
On a toutes et tous vu passer ce genre de post : « Tu te réveilles entre 2h et 3h du matin ? C’est ton foie qui travaille. » Et souvent, la conclusion implicite qui suit : quelque chose ne va pas chez toi.
Avant de décortiquer pourquoi ce type d’affirmation peut devenir problématique, on doit d’abord comprendre d’où elle vient. Car derrière ces posts simplistes se cache un système de pensée cohérent et sophistiqué qui mérite mieux qu’un résumé en deux lignes.
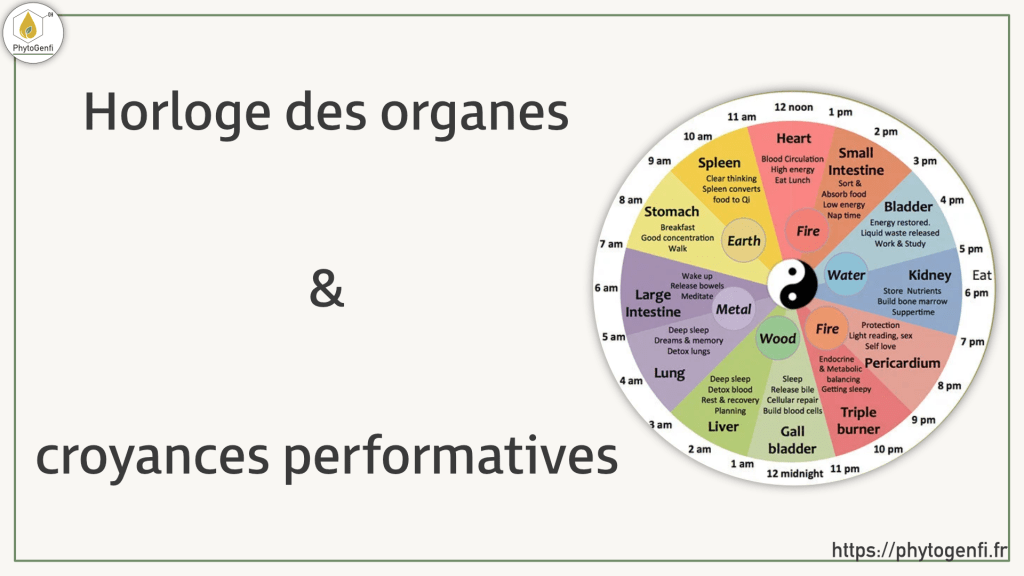
Le cycle Yin/Yang sur 24 heures
En Médecine Traditionnelle Chinoise, tout ce qui existe oscille entre deux forces complémentaires et opposées : le Yin et le Yang. Ce ne sont pas des entités mystérieuses, ce sont des qualités relatives. Le Yang c’est le mouvement, la chaleur, l’activité, l’expansion. Le Yin c’est le repos, la fraîcheur, l’intériorisation, la régénération. Et comme tout dans la nature, ces deux forces suivent un rythme cyclique sur 24 heures.
Le jour est Yang. C’est le temps de l’action, de la dépense énergétique, de l’interaction avec le monde extérieur. La nuit est Yin. C’est le temps du retrait, de la récupération, de la régénération des tissus et des organes.
C’est dans ce cadre global que s’inscrit ce qu’on appelle l’horloge biologique de la MTC. Le Qi, l’énergie vitale, circule en permanence dans le corps à travers un réseau de méridiens. Sur ce cycle de 24 heures, chaque organe connaît une fenêtre de deux heures pendant laquelle son activité est à son maximum. Entre 1h et 3h du matin, c’est le foie qui est à son pic. Entre 3h et 5h, c’est le poumon. Entre 17h et 19h, ce sont les reins.
Un outil de diagnostic, pas une alarme
Et c’est là qu’on peut poser une première question simple. L’horloge des organes s’applique sur 24 heures, jour et nuit. Entre 17h et 19h, ce sont les reins qui sont à leur pic d’activité. Si leur énergie est perturbée, on pourrait s’attendre à ressentir à ce moment-là une fatigue inhabituelle, un manque d’élan, une sensation de vide dans le bas du dos. Ce serait le signal diurne équivalent au réveil nocturne.
Pourtant personne ne consulte pour ça. On attribue cette fatigue de fin de journée au travail, au stress, à la vie. On ne pense pas aux reins.
Ce décalage est révélateur. Ce n’est pas vraiment l’horloge des organes qui inquiète, c’est la nuit. Se réveiller dans l’obscurité active quelque chose de plus archaïque, une vigilance, une vulnérabilité. Et c’est sur ce terrain émotionnel déjà fertile que vient se greffer la croyance « mon foie va mal. »
On n’a pas pathologisé l’horloge, on a pathologisé la nuit.
Il y a par ailleurs une différence fondamentale entre un organe qui est à son pic d’activité et un organe qui dysfonctionne. Dans la logique de l’horloge, le foie qui « travaille » entre 1h et 3h du matin fait exactement ce qu’il est censé faire : filtrer le sang, métaboliser les toxines, préparer l’énergie du lendemain. C’est sa fenêtre de régénération Yin, précisément parce qu’on est au repos.
Un symptôme qui apparaît de façon récurrente à cette heure, comme des crampes, une douleur au flanc droit, une sensation de chaleur persistante, c’est différent. C’est un signal qui mérite attention dans le cadre d’un bilan global. Mais un simple réveil ? C’est autre chose.
Pour un praticien en MTC sérieusement formé, un réveil à 3h du matin n’est qu’un indice parmi une centaine d’autres. Il observera la langue, prendra les pouls, évaluera le teint, interrogera l’état émotionnel, l’alimentation, le contexte de vie. Isoler l’horaire du réveil pour en faire un diagnostic, c’est comme regarder uniquement le voyant d’essence pour évaluer l’état mécanique d’une voiture. C’est réducteur, et souvent tout simplement faux.
Heure solaire ou heure officielle ?
Une question qui vient souvent avec : l’horloge des organes se lit-elle sur l’heure officielle ou sur l’heure solaire ? En France par exemple, l’heure légale peut être décalée d’une à deux heures par rapport au soleil selon la saison. Alors si on se réveille à 3h du matin, est-on vraiment dans la fenêtre du foie ?
La vraie réponse c’est que cette question n’a plus vraiment de sens aujourd’hui, et comprendre pourquoi est plus instructif que d’y répondre directement.
L’horloge des organes a été élaborée sur des populations soumises au rythme naturel du soleil. Le lever du jour déclenchait l’éveil, le coucher du soleil annonçait le repos. Le cycle Yin/Yang s’incarnait littéralement dans l’alternance naturelle lumière/obscurité. C’est cette alternance qui calait l’horloge biologique de chacun.
Or on vit aujourd’hui dans un monde de lumière artificielle permanente. Nos soirées sont éclairées, nos écrans émettent une lumière bleue qui leurre notre cerveau en lui faisant croire qu’il fait encore jour. Notre horloge interne, ce qu’on appelle le rythme circadien, est chroniquement perturbée. Elle ne se synchronise plus aussi fidèlement sur le soleil.
Ce qui veut dire que deux personnes qui se couchent à des heures très différentes, qui travaillent de nuit, qui voyagent régulièrement, auront des cycles biologiques décalés l’un par rapport à l’autre. Leur « 3h du matin physiologique » n’est pas la même.
Dans ce contexte, vouloir appliquer une grille horaire fixe à des organismes dont les rythmes circadiens sont tous légèrement différents, c’est utiliser un instrument de précision sans avoir étalonné la mesure. Avant de se demander quelle heure lire, il faudrait d’abord comprendre le rythme circadien propre à chaque personne. Et ça, aucun post Instagram ne peut le faire à notre place.
Ce que la physiologie du sommeil nous dit
Posons-nous maintenant une question simple : est-ce qu’il existe une explication physiologique parfaitement banale au fait de se réveiller entre 2h et 3h du matin ?
La réponse est oui.
Notre sommeil est structuré en cycles d’environ 90 minutes, alternant phases de sommeil profond et sommeil paradoxal. À la fin de chaque cycle, on remonte naturellement vers un stade de quasi-éveil. C’est un mécanisme archaïque, probablement hérité de l’époque où il fallait vérifier que la grotte était toujours sûre. Si on se couche à 23h, la fin du deuxième cycle tombe précisément… entre 2h et 3h du matin.
Dans des conditions normales, on se rendort en quelques minutes sans même s’en souvenir. Ce micro-réveil est physiologique, universel, et n’a strictement rien de pathologique. Ce n’est pas le foie qui parle. C’est simplement la fin d’un cycle.
Quand la croyance devient le vrai problème
C’est là que les choses deviennent à la fois intéressantes et préoccupantes.
Imaginons deux scénarios. Dans le premier, on se réveille à 3h, on se retourne, on se rendort. Dans le second, on se réveille à 3h avec en tête l’idée que ce réveil signale un problème de foie. On commence à scanner son corps, à repenser au verre de vin du dîner, à s’inquiéter. Le cortisol monte. L’éveil s’installe. On ne se rendort plus.
Ce qu’on vient de décrire c’est une croyance performative : c’est la croyance elle-même qui produit le symptôme qu’elle est censée expliquer. On appelle ça l’effet nocebo, le pendant négatif de l’effet placebo. Et dans ce cas précis, c’est une information mal transmise qui déclenche une insomnie réelle chez quelqu’un qui n’avait au départ aucun problème de sommeil.
Les spécialistes du sommeil ont même nommé ce phénomène : l’orthosomnie. C’est le trouble du sommeil causé par la surveillance obsessionnelle de son propre sommeil. Chercher à tout prix à comprendre pourquoi on se réveille peut suffire à empêcher de se rendormir.
Ce qu’on peut en retenir
Tout cela ne disqualifie pas la MTC ni l’horloge des organes. Un outil de diagnostic élaboré sur des siècles d’observation clinique mérite le respect et l’étude sérieuse. Mais utilisé hors de son contexte, réduit à une grille de lecture simpliste, il peut se retourner contre les personnes qu’il est censé aider.
Et puis posons-nous une dernière question : si notre foie avait réellement un problème, est-ce qu’un simple réveil nocturne serait le seul signal ? La réponse est clairement non. Un foie qui souffre s’exprime de bien d’autres façons : une fatigue persistante au réveil malgré une nuit suffisante, des troubles digestifs, une peau qui change de teinte, des urines foncées, une sensibilité accrue à l’alcool ou aux graisses, parfois une gêne sous les côtes droites. Ce sont ces signaux combinés, dans leur cohérence et leur persistance, qui méritent attention. Pas un réveil isolé à 3h du matin.
La prochaine fois qu’on nous dit que notre réveil de 3h signale un problème de foie, on peut se rappeler deux choses. D’abord que c’est peut-être simplement la fin d’un cycle de sommeil, un phénomène aussi naturel que de respirer. Et ensuite que si on commence à s’en inquiéter, c’est cette inquiétude, et non le foie, qui risque de nous empêcher de nous rendormir.
Récemment avec Kévin on s’interrogeait sur ce qui semble être un paradoxe. Parmi les plantes médicinales, l’échinacée occupe une place singulière. Si elle est plébiscitée pour prévenir le rhume, elle fait l’objet d’une mise en garde sévère en Europe dès lors qu’une pathologie auto-immune est déclarée. Pourtant, outre-Atlantique, la vision est tout autre et peut être conseillée en cas de maladie auto-immune. Pourquoi un tel fossé entre ces deux approches ? La réponse ne se trouve pas dans une erreur médicale, mais dans l’espèce, la partie utilisée,la chimie et la manière dont nous transformons la plante.
I. Le dogme européen : Le principe de précaution
En Europe, les instances de santé, notamment l’EMA, classent les Échinacées parmi les plantes utilisées pour prévenir et accompagner les rhumes. Elles sont décrites comme soutenant la réponse immunitaire, en particulier lors des infections respiratoires hautes.
Cependant, dans les pathologies auto-immunes, où la réponse immunitaire est déjà dysrégulée et dirigée contre le soi, renforcer ou activer certaines voies immunitaires peut poser question. L’usage des échinacées repose donc sur un raisonnement de prudence, afin d’éviter d’amplifier une inflammation déjà excessive.
Les fractions polysaccharidiques et glycoprotéiques hydrosolubles, abondantes dans les préparations fraîches d’Échinacée pourpre (E. purpurea), activent les macrophages via les voies de l’immunité innée, augmentant la phagocytose et la sécrétion de médiateurs pro-inflammatoires comme le TNF-alpha et l’IL-1. Cette activation non spécifique de la réponse inflammatoire explique les réserves émises en contexte auto-immun.
II. L’approche anglo-saxonne : La modulation par les récepteurs CB2
En Amérique du Nord, le regard porté sur l’Échinacée est sensiblement différent. Les herboristes cliniciens l’utilisent fréquemment comme un immunomodulateur, avec l’objectif non pas de « stimuler » le système immunitaire, mais de le recalibrer lorsque celui-ci est dysrégulé.
Ce point de vue repose sur l’étude d’une autre famille de composés bioactifs : les alkylamides. Particulièrement abondants dans les racines d’Echinacea angustifolia, espèce sauvage d’Amérique du Nord historiquement privilégiée aux États-Unis, ces composés lipophiles présentent une bonne biodisponibilité orale. Leur structure leur permet d’interagir avec les récepteurs CB2 du système endocannabinoïde, largement exprimés à la surface des cellules immunitaires.
Contrairement aux polysaccharides hydrosolubles, les alkylamides exercent une action régulatrice sur la réponse immunitaire. Ils sont capables d’inhiber une production excessive de cytokines pro-inflammatoires, notamment le TNF-alpha, tout en préservant les mécanismes de surveillance immunitaire vis-à-vis des agents pathogènes. Cette modulation fine explique pourquoi certaines pratiques nord-américaines envisagent l’usage de l’Échinacée, sous conditions strictes d’espèce, de partie utilisée et de forme galénique, y compris dans des contextes où l’immunité est déséquilibrée.
III. Biotope et galénique
En Europe, les préparations sont le plus souvent issues de cultures d’Echinacea purpurea en climat tempéré, transformées en jus pressé ou en extraits aqueux. Ces formes galéniques concentrent principalement les fractions hydrosolubles, notamment les polysaccharides et les glycoprotéines, historiquement associées à une activation de l’immunité innée.
En Amérique du Nord, l’usage traditionnel privilégie davantage les racines d’E. angustifolia, parfois issues de récoltes sauvages, transformées en teintures hydroalcooliques à fort degré. Ces procédés extraient préférentiellement les composés lipophiles, en particulier les alkylamides, dont l’activité relève davantage d’une modulation des voies inflammatoires via le système endocannabinoïde.
Autrement dit, il ne s’agit pas du même profil phytochimique, ni du même effet biologique.
Ainsi, chez une personne vivant avec une maladie auto-immune, la prudence reste indispensable. Mais il est essentiel de comprendre que toutes les Échinacées ne se valent pas. Une teinture mère de racines riche en alkylamides n’aura pas le même impact immunologique qu’un jus de plante fraîche dominé par les polysaccharides.
En phytothérapie, ce n’est jamais seulement la plante qui compte. C’est l’espèce, l’organe, le solvant d’extraction et donc la chimie réellement administrée.
IV. La controverse de la pandémie (COVID-19)
Cette divergence théorique a pris une tournure très concrète au printemps 2020. En pleine première vague de COVID-19, l’ANSES (Agence nationale de sécurité sanitaire française) a émis une alerte déconseillant formellement l’usage de l’échinacée en automédication.
La raison invoquée ? La crainte de l’orage cytokinique. Les autorités sanitaires, se basant sur la classification européenne « immunostimulante », ont redouté que la plante n’aggrave l’inflammation pulmonaire en augmentant la production de cytokines (IL-6, TNF-alpha), mettant ainsi les patients en danger.
Pourtant, quelques mois plus tard, une étude suisse menée par le laboratoire de Spiez (Signer et al., 2020) venait nuancer cette crainte. En travaillant sur un extrait hydro-alcoolique spécifique (Echinaforce), les chercheurs ont observé in vitro que non seulement l’extrait désactivait le virus, mais qu’il réduisait la libération des cytokines pro-inflammatoires au lieu de les augmenter.
Cet épisode illustre parfaitement le paradoxe : l’autorité sanitaire alertait contre le risque théorique des polysaccharides (stimulation), tandis que la recherche observait l’effet protecteur réel des alkylamides (modulation).
V. Comment décoder un flacon d’échinacée ?
Pour savoir si vous avez affaire à un produit plutôt « stimulant » (Axe Européen) ou « modulateur » (Axe Anglo-saxon), l’étiquette est votre meilleure alliée. Voici les trois indicateurs clés à surveiller :
1. La partie de la plante utilisée
C’est l’indice le plus simple.
Parties aériennes / Jus de plante fraîche : Très riches en polysaccharides. C’est la forme « classique » européenne (ex: Echinaforce ou jus stabilisés). Usage : Prévention robuste chez le sujet sain, mais prudence en cas d’auto-immunité.
Racines (Radix) : C’est là que se concentrent les précieux alkylamides. Un produit à base de racines est généralement plus orienté vers la modulation.
2. Le mode d’extraction (le solvant)
Le solvant choisit les molécules qu’il emporte avec lui.
Extrait aqueux / Jus pressé / Infusion : L’eau n’extrait pas les alkylamides. Ces produits contiennent essentiellement des polysaccharides. (Risque de stimulation brute).
Teinture / Extrait hydro-alcoolique : L’alcool est indispensable pour extraire les alkylamides (lipophiles). Plus le degré alcoolique est élevé (entre 45° et 65°), plus le produit sera riche en composés modulateurs agissant sur les récepteurs CB2.
3. Les indicateurs de qualité : Le « tingle factor »
Les alkylamides ont une propriété physique unique : ils provoquent un picotement ou un engourdissement caractéristique sur la langue (similaire au poivre du Sichuan).
Si votre teinture d’échinacée ne produit aucun picotement après quelques secondes en bouche, elle est probablement pauvre en alkylamides et ne jouera pas son rôle de modulateur.
Tableau récapitulatif pour l’étiquetage
Ce que vous lisez sur l’étiquette
Profil dominant
Action probable
« Jus de plante fraîche pressée »
Polysaccharides
Stimulation (Axe Européen)
« Extrait de racines (alcool 60%) »
Alkylamides
Modulation (Axe Anglo-saxon)
« Standardisé à X % d’échinacosides »
Composés phénoliques
Antioxydant / Antibactérien
« Poudre de plante totale (gélules) »
Mixte (faible dosage)
Action douce et lente
VI. Références et preuves scientifiques
La recherche récente commence à valider cette nuance. Une étude marquante de Gertsch et al. (2004) a démontré que les alkylamides de l’échinacée agissent comme des ligands des récepteurs CB2, expliquant leur effet immunomodulateur plutôt que simplement stimulant.
Par ailleurs, une étude clinique sur l’extrait Echinaforce (riche en alkylamides) a montré une réduction des cytokines inflammatoires lors d’infections, suggérant qu’un extrait bien standardisé ne provoque pas la redoutée « poussée » auto-immune, mais aide au contraire à la résolution de l’inflammation (Ritchie et al., 2011).
Sources sélectionnées pour approfondir :
Gertsch J, et al. (2004).Echinacea alkylamides modulate TNF-alpha gene expression via cannabinoid receptor type 2 (CB2). FEBS Letters.
Ritchie M, et al. (2011).Effects of Echinaforce® on ex vivo cytokine secretion by human peripheral blood mononuclear cells. Phytomedicine.
Boon H, & Smith M. (2009).The Complete Natural Medicine Guide to the 50 Most Common Medicinal Herbs. (Sur la différence de pratique clinique US/Canada).
Raduner S, et al. (2006).Alkylamides from Echinacea Are a New Class of Cannabinomimetics. Journal of Biological Chemistry.
Signer J, et al. (2020).In vitro virucidal activity of Echinaforce®, an Echinacea purpurea preparation, against coronaviruses, including common cold coronavirus 229E and SARS-CoV-2. Virology Journal. (L’étude clé sur la période COVID)..
Cet article présente une revue critique de la littérature concernant l’impact de la diffusion d’huiles essentielles (HE) sur la qualité de l’air intérieur (QAI) et la santé humaine. L’objectif est d’évaluer la sécurité des différents protocoles de diffusion en analysant les émissions chimiques, les polluants secondaires et les effets biologiques rapportés.
Typologie des dispositifs et émissions
Les études analysées distinguent quatre catégories principales de diffuseurs, chacune présentant un profil d’émission distinct :
Diffuseurs passifs (capillarité, tiges) : Ces dispositifs émettent des composés organiques volatils (COV) à des taux faibles mais constants (environ 4 mg/h pour le γ-terpinène). L’impact sur la QAI est durable, pouvant s’étendre de 5 heures à 60 jours selon la charge d’huile. Angulo-Milhem et al. (2021) notent que les espèces volatiles comme l’α-pinène sont émises rapidement, tandis que d’autres persistent plusieurs semaines.
Diffuseurs ultrasoniques (brumisateurs) : Ils génèrent des masses importantes de COV (ex: ~2,6 mg de d-limonène par essai). Le point critique concerne les émissions particulaires : l’utilisation d’eau du robinet entraîne une production massive de particules fines (PM10 ~562 µg/m³, PM2.5 ~452 µg/m³) due aux minéraux dissous, dépassant largement les seuils sanitaires7. L’eau distillée réduit considérablement ces émissions.
Diffuseurs par nébulisation (Venturi) : Ces appareils produisent les pics d’émission les plus élevés (ex: 66-82 mg/h/g pour le γ-terpinène), entraînant des concentrations transitoires très fortes de terpènes dans l’air.
Diffuseurs à chaleur (bougies, brûleurs) : Ils provoquent des pics de COV rapides (en 30-60 min) et, dans le cas des bougies, émettent des sous-produits de combustion comme le CO et le CO2. Su et al. (2007) rapportent des concentrations élevées de linalol (jusqu’à 787 µg/m³) avec ces dispositifs.
Impact physico-chimique sur la qualité de l’air
La diffusion d’HE introduit des polluants primaires et favorise la formation de polluants secondaires.
Composés organiques volatils (COV)
Tous les modes de diffusion augmentent la charge en COV (terpènes, limonène, alpha-pinène). Dans des scénarios réalistes, les concentrations chroniques moyennes peuvent atteindre des niveaux préoccupants (jusqu’à 8 mg/m³ pour le limonène dans les pires cas).
Chimie de l’air et polluants secondaires
La réaction des terpènes avec l’ozone ambiant (O₃) est une source majeure de préoccupation. Palmisani et al. (2020) ont montré que cette réaction génère rapidement :
Des aldéhydes irritants comme le formaldéhyde, l’acétone et l’acétaldéhyde.
Des particules ultrafines et des aérosols organiques secondaires (AOS), avec des pics de nombre de particules atteignant 5×10⁴ part/cm³.
Matières particulaires (PM)
Outre les particules issues de la chimie secondaire, les diffuseurs ultrasoniques utilisant de l’eau du robinet sont une source directe et massive de particules fines (PM2.5 et nanoparticules), indépendamment de la chimie des gaz. Rimdjonoka et al. (2024) rapportent des niveaux de PM2.5 supérieurs à 450 µg/m³ lors de sessions de 4 heures avec de l’eau du robinet.
Effets sur la santé et populations vulnérables
Irritation et toxicité respiratoire
L’effet le plus fréquemment observé est l’irritation sensorielle des yeux et des voies respiratoires supérieures. Wolkoff (2020) souligne que cette irritation est souvent due aux produits d’oxydation (formaldéhyde, 4-OPA) plutôt qu’aux terpènes eux-mêmes. Bien que l’inflammation manifeste des voies respiratoires soit rare aux niveaux domestiques, des réponses de stress physiologique sont observées.
Groupes à Risque
Les enfants, les personnes âgées, les femmes enceintes et les asthmatiques sont identifiés comme vulnérables.
Asthmatiques : Ils peuvent percevoir moins l’irritation sensorielle que les sujets sains, mais présenter des réponses de stress physiologique (augmentation de l’α-amylase salivaire) plus importantes.
Sensibilisation : Les constituants des HE peuvent agir comme sensibilisants cutanés et contribuer aux symptômes respiratoires.
Conclusions de sécurité et recommandations
L’analyse des données permet de classer la sécurité des dispositifs selon les conditions d’utilisation :
Sûr sous condition :
Diffuseurs passifs : Si la charge d’huile est faible et la ventilation maintenue.
Ultrasoniques :Uniquement avec de l’eau distillée/déminéralisée, sessions courtes (<30 min) et bonne ventilation.
Nébuliseurs : Uniquement en usage intermittent et faible dose.
Potentiellement nocif :
Ultrasoniques avec eau du robinet : En raison des émissions massives de particules.
Chaleur (bougies) : En raison des pics de COV et de la combustion.
Sprays : Risque élevé de dépassement des seuils aigus d’exposition.
Recommandations pratiques
Ventilation : Assurer un taux de renouvellement d’air ≥ 1 h⁻¹ et ventiler 30 à 60 minutes après diffusion.
Eau : Toujours utiliser de l’eau distillée ou déminéralisée pour les diffuseurs ultrasoniques.
Ozone : Éviter la diffusion lors des pics d’ozone ou en présence d’appareils générant de l’ozone.
Vulnérabilité : Éviter la diffusion en présence de personnes asthmatiques, enceintes ou sensibles.
Bibliographie
Angulo-Milhem, S., Verriele, M., Nicolas, M., & Thevenet, F. (2021). Indoor use of essential oils: Emission rates, exposure time and impact on air quality. Atmospheric Environment, 244, 117863. https://doi.org/10.1016/j.atmosenv.2020.117863
Du, B. (2022). The Impact of Indoor Air Quality and Exposure to Indoor Air Pollutants on Cognitive Function. (Données extraites sur les diffuseurs ultrasoniques).
Karr, G., Quivet, E., Ramel, M., & Nicolas, M. (2022). Sprays and diffusers as indoor air fresheners: Exposure and health risk assessment based on measurements under realistic indoor conditions. Indoor Air, 32. https://doi.org/10.1111/ina.12923
Kim, S., Hong, S.-H., Bong, C.-K., & Cho, M.-H. (2015). Characterization of air freshener emission: the potential health effects. The Journal of Toxicological Sciences, 40(5), 535-550. https://doi.org/10.2131/jts.40.535
Palmisani, J., Nørgaard, A. W., Kofoed-Sørensen, V., Clausen, P. A., de Gennaro, G., & Wolkoff, P. (2020). Formation of ozone-initiated VOCs and secondary organic aerosol following application of a carpet deodorizer. Atmospheric Environment, 222, 117149. https://doi.org/10.1016/j.atmosenv.2019.117149
Rimdjonoka, R., Veignere, E., Laicāns, I., Martinsone, Ž., & Skadiņš, I. (2024). The effect of ultrasonic diffuser on indoor air quality. SOCIETY. INTEGRATION. EDUCATION. Proceedings of the International Scientific Conference, 2, 713-722. https://doi.org/10.17770/sie2024vol2.7837
Su, H.-J., Chao, C.-J., Chang, H.-Y., & Wu, P.-C. (2007). The effects of evaporating essential oils on indoor air quality. Atmospheric Environment, 41, 1230-1236. https://doi.org/10.1016/j.atmosenv.2006.09.044
Wolkoff, P. (2020). Indoor air chemistry: Terpene reaction products and airway effects. International Journal of Hygiene and Environmental Health, 225, 113439. https://doi.org/10.1016/j.ijheh.2019.113439
Le clou de girofle est utilisé depuis des siècles pour calmer les rages de dents, mais ce n’est que récemment que la pharmacologie moléculaire a permis de comprendre précisément pourquoi son principe actif, l’eugénol, est si efficace.
Loin d’être un simple placebo sensoriel, l’eugénol est un agent pharmacologique complexe qui agit comme un « anesthésique sale » (dans le sens noble du terme scientifique) : il ne cible pas un seul mécanisme, mais verrouille le système nerveux par trois voies distinctes simultanées.
L’eugénol agit sur trois fronts
Pour qu’une douleur dentaire soit perçue, le nerf doit générer un signal électrique, le transmettre, et le relayer chimiquement de neurone en neurone jusqu’au cerveau. L’eugénol intervient à chacune de ces étapes.
1. Blocage des canaux sodiques (Nav)
C’est le mécanisme principal, comparable à celui de la lidocaïne utilisée par les dentistes.
Le fonctionnement normal : L’influx nerveux est une vague électrique créée par l’entrée massive d’ions Sodium (Na+) dans la fibre nerveuse via des canaux voltage-dépendants.
L’action de l’eugénol : La molécule vient se loger physiquement à l’intérieur du canal, bloquant le passage.
La spécificité : Contrairement à certains anesthésiques qui ne bloquent que les canaux ouverts (actifs), l’eugénol semble capable de se lier aux canaux quel que soit leur état. Sans entrée de sodium, la dépolarisation est impossible : le « câble » est coupé, le message de douleur ne part pas.
2. Désensibilisation des récepteurs TRP
Ce mécanisme explique la sensation paradoxale de brûlure suivie d’un engourdissement.
L’activation initiale : L’eugénol est un agoniste des récepteurs TRPV1 (sensibles à la chaleur et à l’acidité). Au moment de l’application, il les active violemment, créant un picotement.
Le crash du système : Cette sur-stimulation entraîne une désensibilisation rapide. Le récepteur, saturé, se « verrouille » et devient réfractaire. Il ne répond plus, ni à l’eugénol, ni aux stimuli inflammatoires de la dent.
3. Inhibition des canaux calciques (Ca2+)
Le rôle du calcium : En bout de course, pour transmettre l’information au neurone suivant, la terminaison nerveuse doit laisser entrer du calcium. Ce calcium déclenche l’éjection des neurotransmetteurs de la douleur (comme la Substance P ou le glutamate).
L’action de l’eugénol : En inhibant les courants calciques à haut voltage (HVA), l’eugénol empêche la libération de ces messagers chimiques. Même si un signal résiduel arrivait, il ne pourrait pas franchir la synapse.
II. Le secret de la structure Phénol-Méthoxy
Si l’eugénol est un anesthésiant puissant, pourquoi le thymol ou le carvacrol , qui sont aussi des phénols, ne le sont-ils pas autant ? La réponse réside dans la géométrie moléculaire.
Le problème du Thymol et du Carvacrol
Ces molécules possèdent une fonction phénol (un anneau benzénique avec un groupe -OH) et une chaîne carbonée (isopropyle).
Elles sont très lipophiles (grasses), ce qui leur permet d’entrer dans la membrane du nerf.
Mais pour se fixer sur le récepteur protéique (le canal sodique), elles ne disposent que d’un seul « point d’accroche » : le groupe -OH. La liaison est instable. De plus, leur groupe isopropyle est volumineux, créant une gêne stérique qui empêche un ajustement parfait dans le canal.
L’effet « Pince » de l’eugénol
L’eugénol possède une particularité structurelle décisive : une fonction méthoxy (-OCH3) placée en position ortho (juste à côté) du groupe phénol.
Une liaison bidentate (à deux dents) : Grâce à la proximité du groupe -OH et du groupe -OCH3, l’eugénol peut former deux liaisons simultanées avec la protéine du canal :
Le -OH donne une liaison hydrogène.
L’oxygène du -OCH3 accepte une liaison hydrogène. Ça ancre solidement la molécule dans le site de liaison, là où le thymol « glisse ».
L’équilibre parfait : Le groupe méthoxy apporte une légère polarité qui manque aux autres phénols trop gras. Cet équilibre hydrophile/lipophile permet à l’eugénol de naviguer idéalement entre la membrane grasse du neurone et le cœur aqueux du canal ionique.
L’efficacité de l’eugénol n’est pas un hasard. C’est le résultat d’une synergie parfaite entre une capacité à bloquer de multiples voies de la douleur (Sodium, TRP, Calcium) et une architecture chimique spécifique (Phénol + Méthoxy) qui lui assure une affinité supérieure pour les cibles nerveuses par rapport aux autres molécules aromatiques.
Références scientifiques
Park, C. K., Kim, K., Jung, S. J., Kim, M. J., Ahn, D. K., Hong, S. D., … & Oh, S. B. (2006). Eugenol inhibits sodium currents in dental afferent neurons. Journal of Dental Research, 85(10), 900-904.
Lee, M. H., Yeon, K. Y., Park, C. K., Li, H. Y., Fang, Z., Kim, M. S., … & Oh, S. B. (2005). Eugenol inhibits calcium currents in dental afferent neurons. Journal of Dental Research, 84(9), 848-851.
Yang, B. H., Piao, Z. G., Kim, Y. B., Lee, C. H., Lee, J. K., Park, K., … & Oh, S. B. (2003). Activation of vanilloid receptor 1 (VR1) by eugenol. Journal of Dental Research, 82(10), 781-785.
Kamatou, G. P., Vermaak, I., & Viljoen, A. M. (2012). Eugenol—from the remote Maluku Islands to the international market: an ethno-pharmacological and phytochemical review. Molecules, 17(6), 6953-6981.
Reuter, S., Fortin, H., Vidari, G., & Just, M. J. (2008). Anesthetic activity of essential oils and their components. Planta Medica, 74(09), PA93.